基因组重复序列分类 转座子
真核生物基因组重复序列分类:
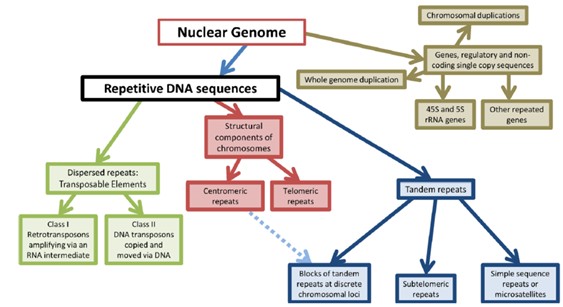 研究最多的是散在重复序列,也就是transposon
研究最多的是散在重复序列,也就是transposon
转座子(transposon,Tn)是真核生物基因组中普遍存在的可移动和自主复制的DNA重复序列,1950年由著名遗传学家McClintock首先在玉米中发现[1]。根据转座机制不同,转座子可分为反转座子和DNA转座子两大类型。以前认为转座子是无功能的“垃圾DNA”,但目前研究表明,转座子在植物基因组进化、基因表达调控、系统发育和遗传多样性评价方面具有重要作用。
分离、鉴定植物转座子并深入研究其基因调控功能,对利用生物技术开展作物遗传改良具有较高的应用价值。如淀粉合成酶(granule-bound starch synthase,GBSSI)基因决定植物中直链淀粉的合成,转座子通过诱发玉米GBSSI基因突变形成等位基因,从而改良了淀粉品质[2]。然而,对于转座子是如何调控植物基因表达这一问题认识还较为模糊。近年来,国内外学者在植物转座子类型和结构特征、活性及其影响因素、预测方法及标记开发等方面取得一系列成果,跟踪和分析植物转座子的最新研究进展对全面了解转座子的功能具有重要意义。为了理清植物转座子与基因表达调控关系,本文综述了植物转座子分类、起源和转座机制以及转座子与宿主基因组间的表观遗传互作,阐述了不同转座子对基因表达调控方式,并对今后研究前景进行了展望。
1 植物转座子
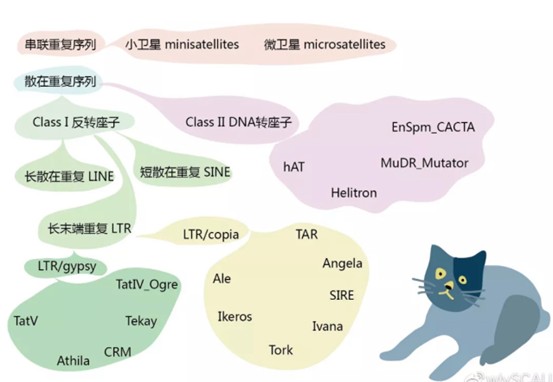
根据转座机制不同,转座子可分为Class Ⅰ元件(反转座子)和Class Ⅱ元件(DNA转座子)两大类型。转座子转座引起插入突变、产生新的基因、产生染色体畸变、引起生物进化,从而引发许多遗传变异。
反转座子(retrotransposon)是一类以RNA为中间媒介,通过“复制-粘贴”方式转座的多拷贝、高度异质性的移动元件。根据是否含有长末端重复序列(long terminal repeat,LTR)可分为LTR-反转座子和非LTR-反转座子。其中,LTR-反转座子主要分为Ty1-copia和Ty3-gypsy两个亚家族,非LTR-反转座子则包括长散布重复序列(long interspersed repetitive element,LINE)和短散布重复序列(short interspersed repetitive element,SINE)两类,LINE负责SINE的转座。反转座子不仅可以在世代中纵向传递,也可在物种间进行水平转移,或者逆转录错误,从而产生高度异质性。在芸薹属近缘种中,转座子的扩增差异造成甘蓝基因组比白菜基因组大,但白菜基因组中转座子插入基因内部频率要高于甘蓝基因组,完整的LTR比单独的LTR具有更强的读出活性,DNA转座子更偏向于插入或保留在基因的内含子中[3]。
DNA转座子(DNA transposon)是通过DNA中间体,以“剪切-粘贴”方式进行转座的单拷贝的转座元件。根据能否自主转座,又可分为自主型DNA转座子和非自主DNA转座子,后者只有在自主型DNA转座子存在时才能进行转座,而前者自身可以编码转座酶进行转座。微小倒置重复转座元件(miniature inverted repeat transposable element,MITE)是一类具有反转座子活性的高拷贝的非自主DNA转座子,1992年,由Bureau等在玉米中发现[4],被认为是基因组进化的驱动力,其数量与植物基因组大小呈显著相关。如玉米基因组的Ac/Ds和Spm/En转座系统[5]。
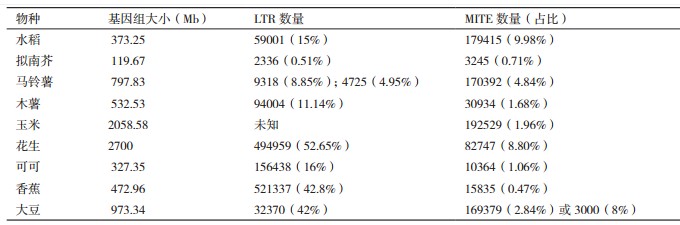
根据转座子的转座酶氨基酸序列差异,高等植物转座子超家族包括LTR反转座子、hAT、CACTA因子、Mutator和MULEs、Tc1/mariner、MITEs等[6]。此外,转座子也可分为插入序列(insertional sequence,IS)、复合转座子和复杂转座子。一些常见植物基因组大小、不同类型转座子数量,见表 1。
Chromosome Res. 2015 Sep;23(3):415-20. doi: 10.1007/s10577-015-9499-z. PMID: 26514350.
何虎翼, 谭冠宁, 唐洲萍, 杨鑫, 李丽淑, 何新民. 植物转座子与基因表达调控[J]. 生物技术通报, 2017, 33(4): 38-43
转座子讲解视频:
https://dls.ym.edu.tw/course/hb/lec07-chromosome-and-genome.html
https://www.youtube.com/playlist?list=PLubKl2ZLAkpp-LLpS1SUbXjLR7LZf0I9E
- 发表于 2022-09-06 10:27
- 阅读 ( 8818 )
- 分类:基因组学
