转录组如何与其他实验手段结合?(第二篇文献)
上一期,我们一起了解了积雪草苷合成过程中糖基化反应的转录组文章,今天我们来看第二篇同样以这个转录组数据为基础的第二篇SCI文章,这一次作者关注点更加细致,下面听我娓娓道来:积雪草酸是由香树脂醇α-amyrin在C-2α, C-23 和C-28三个位置进行3次羟基化形成的。(红色框内)
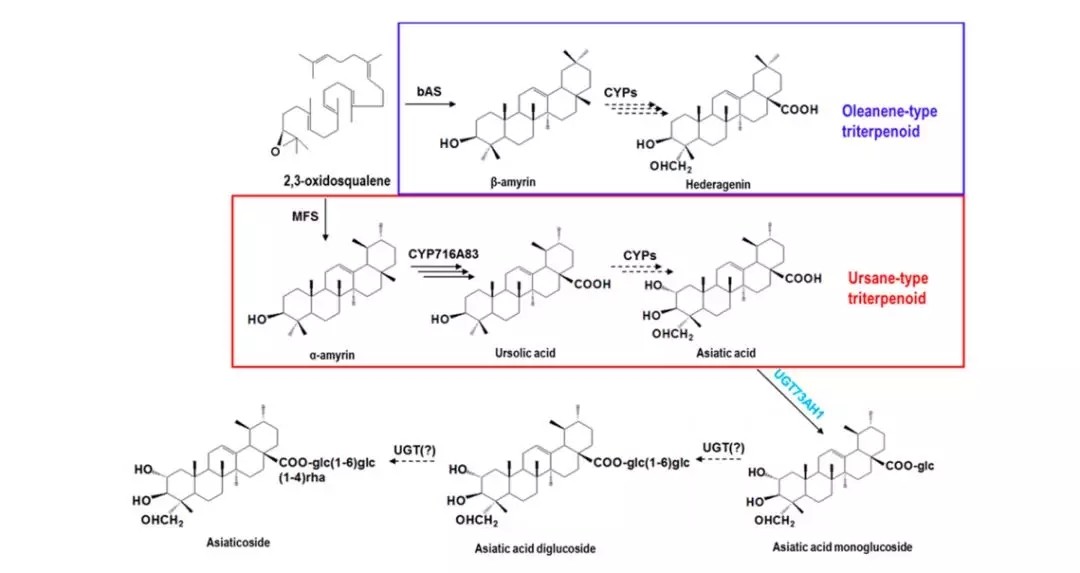
前人已经报道细胞色素P450亚家族CYP716中CYP716A83和CYP716A86单氧酶催化了C-28位置的羟基化,CYP716C11负责C-2α位置的羟基化,而C-23位置则无报道,所以本文重点阐释C-2α位置的羟基化问题。同样作者在该文章依然借助了qRT-PCR、异源共表达、气质联用(GC-MS)等手段,下面一起来看看:
1. 香树脂醇α-amyrin(底物)合成基因CaDDS的验证
由于前人对合成α-amyrin的基因CaDDs报道有争议,所以作者通过在GIL77酵母突变株中过表达CaDDs,并通过质谱鉴定生成物,验证了CaDDs合成α-amyrin的正确性。接下来将用CaDDs作为底物α-amyrin提供者与候选P450单氧酶基因进行共表达试验。
2. p450基因的筛选
作者通过对茉莉酸甲酯处理后的积雪草叶片转录组组装出的unigenes进行同源比对,共筛选出149 unigenes被注释为P450酶基因, 23个具有完整开放阅读框,其中13个在其他物种中报道过与次生代谢有关的被划分到P450家族不同的亚家族。
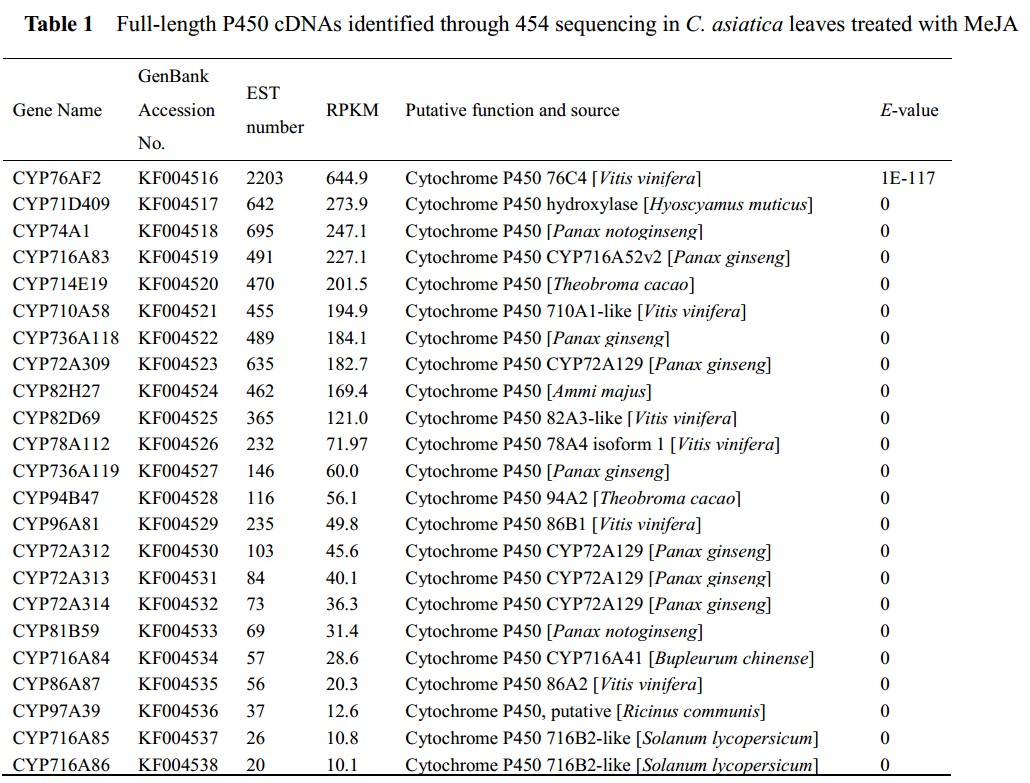
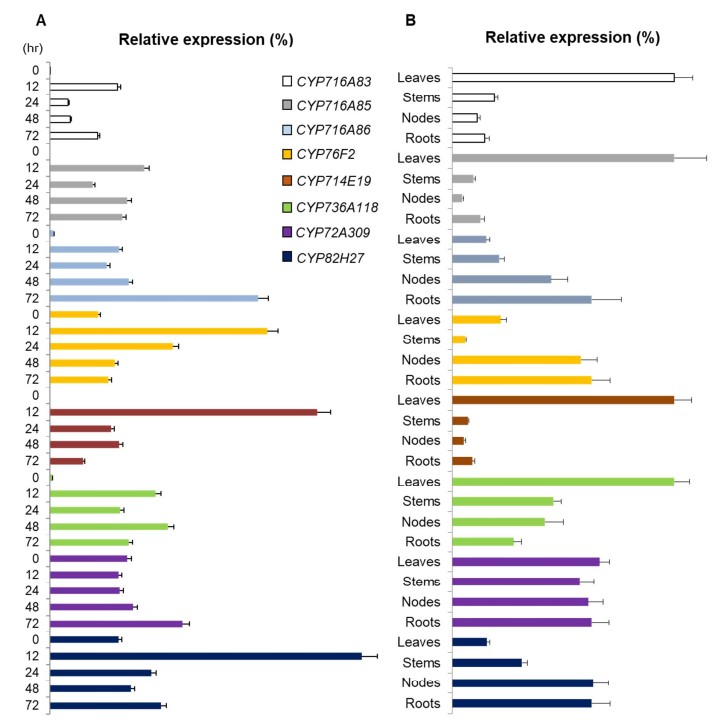
接下来作者又在茉莉酸甲酯处理后的发根培养物中用RT-PCR对上述13个与次生代谢相关的P450酶基因进行表达模式变化分析,结果13个基因中只有8个在处理后12h开始表现出上调表达;进一步用QRT-PCR检测这8个上调表达基因,又排除掉cyp72A309这个酶基因。
由于积雪苷只要是在叶子中合成,根据RNA组织特异性表达原则,作者把在叶中特异表达的P450酶基因作为优先关注的候选基因,最终选定CYP716A83, CYP716A85, CYP714E19 和CYP736A118这4个基因作为P450酶候选基因。
 将积雪草所有的P450酶基因按序列相似性构建进化树,可以看到CYP716A83与已知参与C-28位置的羟基化的P450基因聚在一起;CYP716A85则被重新定义为CYP716E41,参与C-6位置的羟基化;而CYP714E19 和 CYP736A118才是最有可能参与C-23位置羟基化的酶。
将积雪草所有的P450酶基因按序列相似性构建进化树,可以看到CYP716A83与已知参与C-28位置的羟基化的P450基因聚在一起;CYP716A85则被重新定义为CYP716E41,参与C-6位置的羟基化;而CYP714E19 和 CYP736A118才是最有可能参与C-23位置羟基化的酶。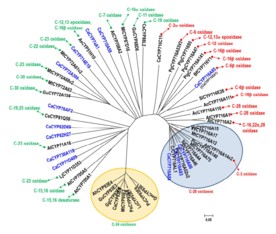
3. 共表达系统检验候选P450酶催化活性
酵母共表达系统检测候选基因酶催化活性,分两轮试验进行,第一轮是单个候选酶基因与CaDDs的共表达;第二轮是多个候选酶基因与CaDDs的共表达,两轮实验产物均以气质联用(GC-MS)进行检测鉴定。
CaDDs与CYP716A83酵母系统共表达,生成物鉴定显示CYP716A83酶能够将多种三萜类包括α/β-amyrin C-28位置甲基分步氧化至羧基,生成三萜类酸。
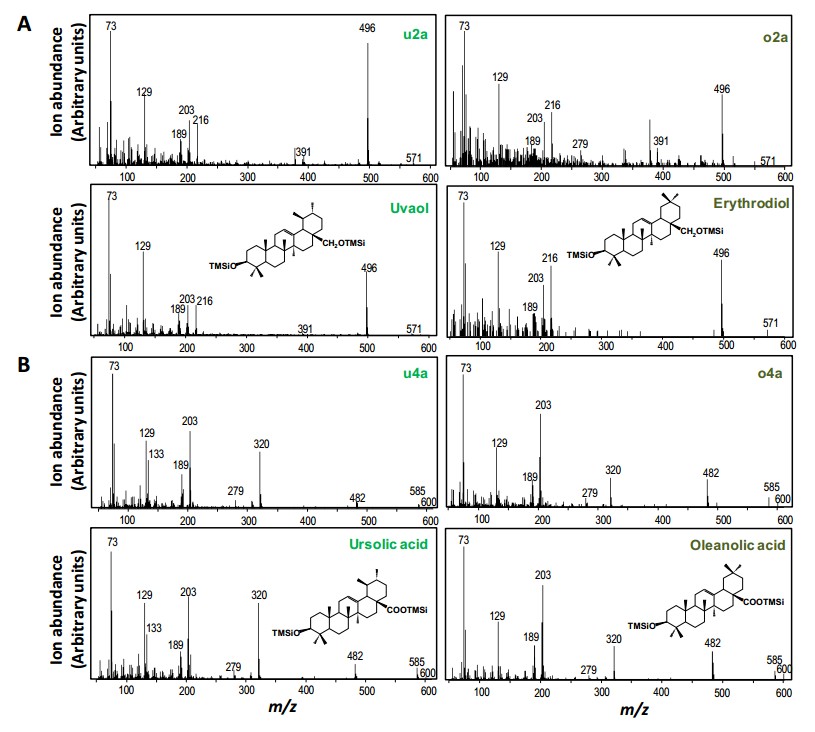
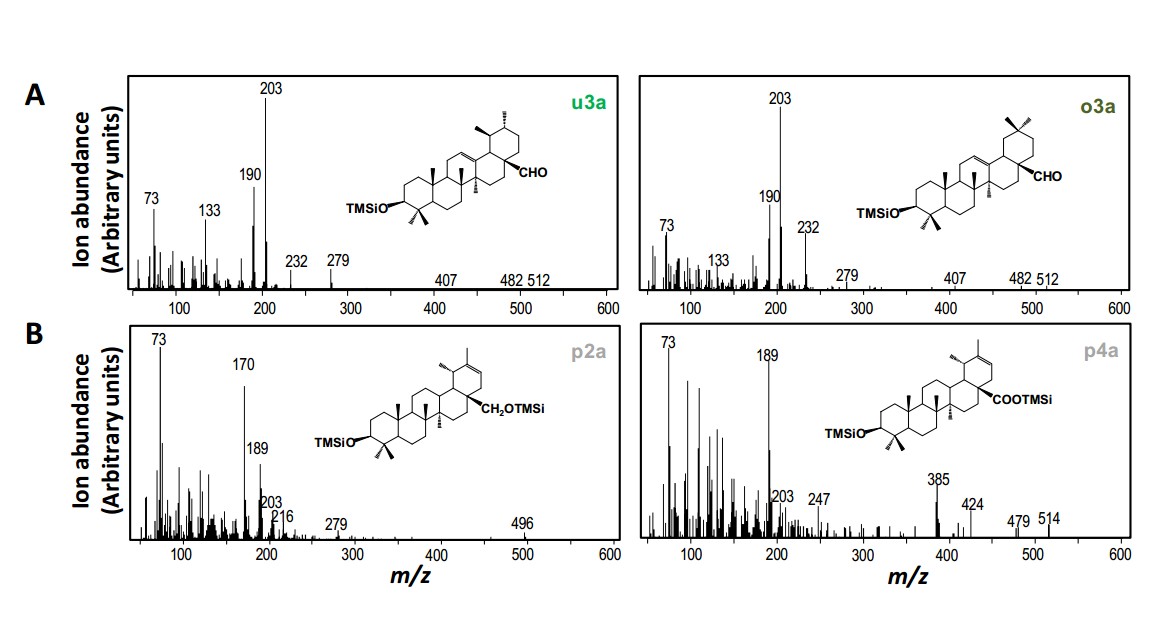
CaDDs分别与其他三个候选P450酶基因酵母共表达体系均能生成三萜醇类,主要是β-amyrin 和α-amyrin,但未发生进一步氧化。
第二轮实验中以CaDDs和CYP716A83分别与CYP716A85、CYP714E19 、 CYP736A118构建共表达体系,结果显示CaDDs、CYP716A83和CYP716A85;CaDDs、CYP716A83和CYP736A118两组合均生成三萜类产物,并未产生进一步的氧化产物。但是CaDDs、CYP716A83和CYP714E19组合可以生成多种C-23被氧化成羟基的三萜类化合物,并且CYP714E19可以催化熊果纯、高根二醇、熊果酸、齐墩果酸,但是不能单独催化香树脂醇(amyrin)。
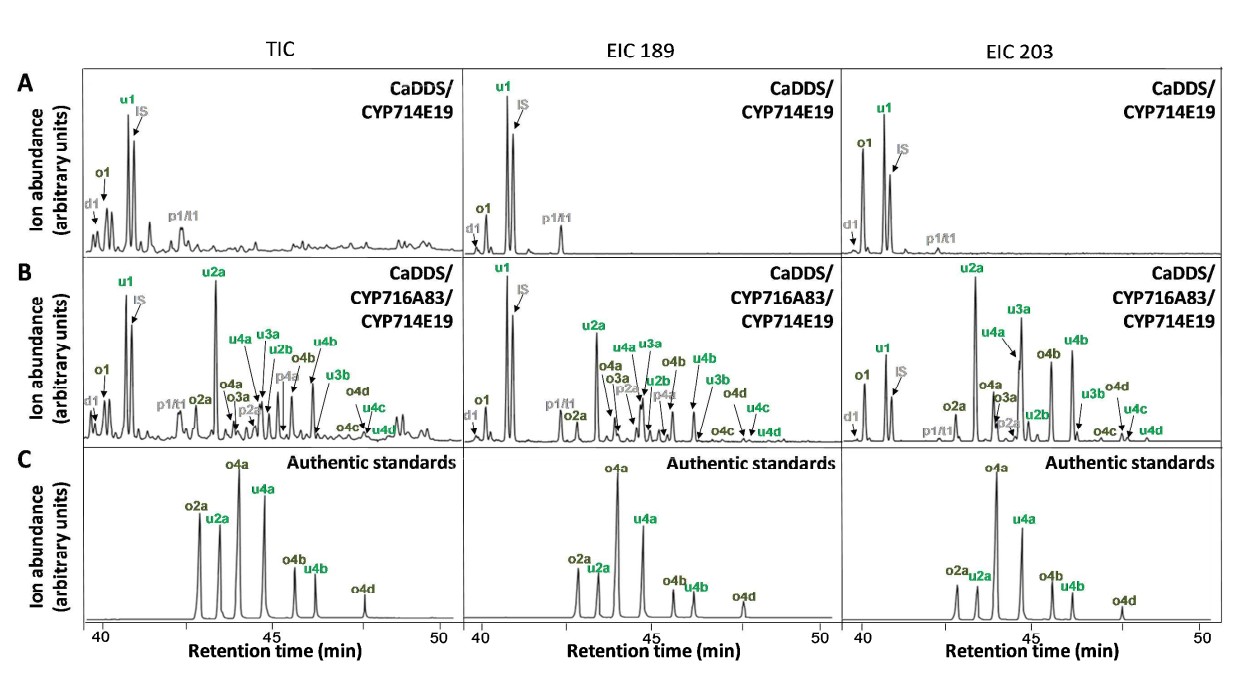
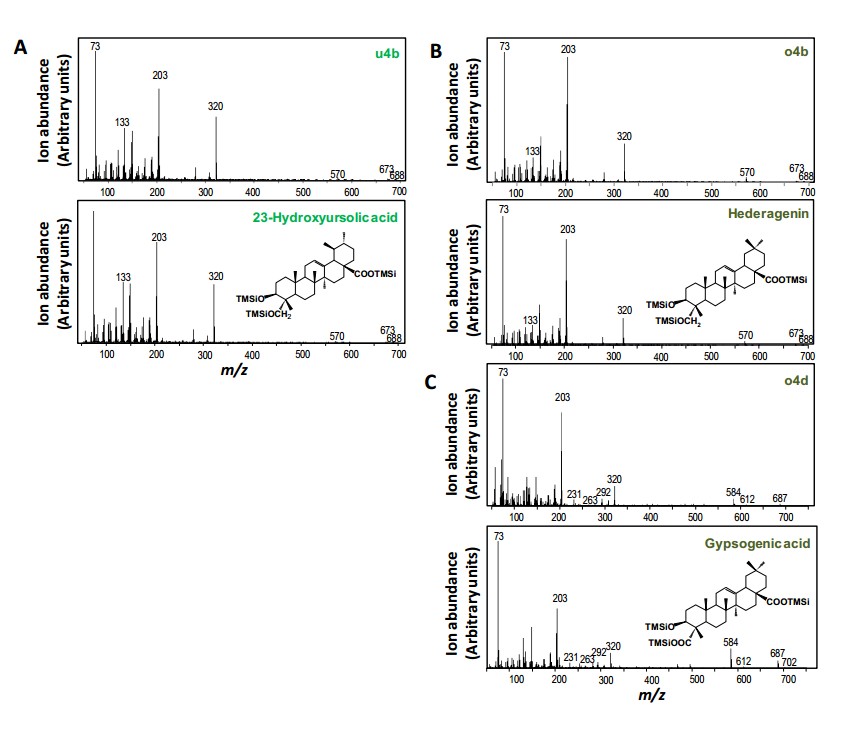
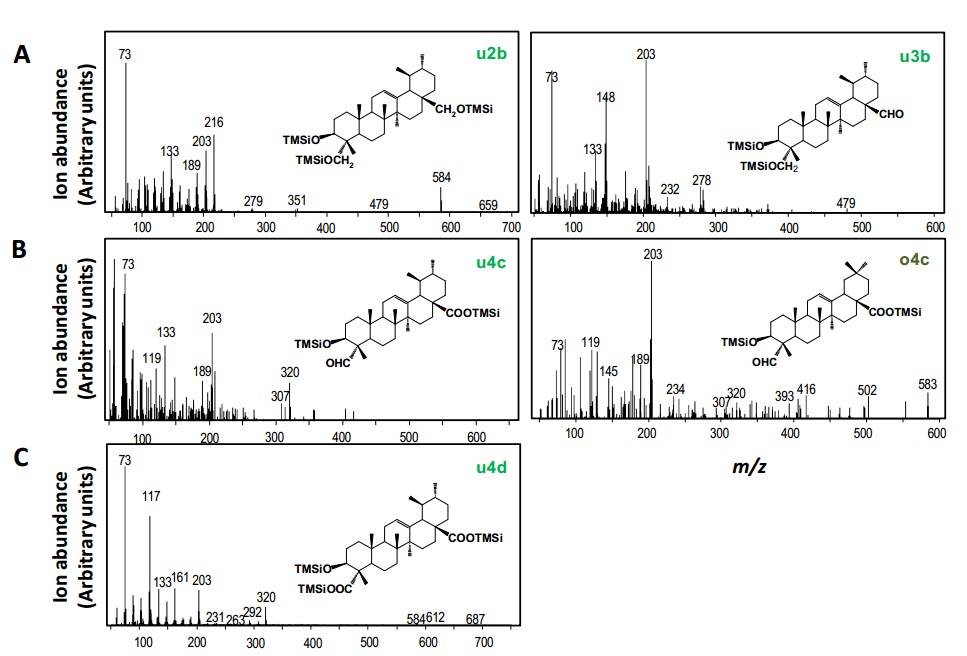
CYP714E19单独不能催化amyrin,但与CYP716A83一起却可以催化amyrin的C-23羟基化,说明有可能CYP716A83催化的C-28位置生成三萜类酸,发生在C-23羟基化之前。
最后作者还提出积雪草中不仅存在乌苏烷类型三萜皂苷,还可能存在齐墩果烷类皂苷,他们共享2α-C, 6β-C和 23-C等功能基团,只是本实验数据的局限性,要想进一步确认还需更多的实验数据支持。

参考文献:
Kim OT at all. A Novel Multifunctional C-23 Oxidase, CYP714E19, Is Involved in Asiaticoside Biosynthesis. Plant Cell Physiol. 2018 Mar 22.
日常科研中你我经会常遇到看不懂的图表,不会挖掘的数据,没有思路的文章,沟通不畅的个性化分析,求人不如求己,一切痛点都能解决:
1. 单细胞/空间转录组正在大火,高分文章必备,0基础学单细胞/空间转录组分析,做更牛的个性化分析,绘更漂亮的图,点这个链接:单细胞/空间转录组分析实操
2. 2区文章发文新思路,0实验就能发文章,省钱省时间,套路化内容好写易发表,0基础学习最新版基因家族分析/泛基因家族分析链接:最新基因家族分析 ;最新版泛基因家族分析
3. 转录组越做越普遍,实验必备,看不懂结果?不会深入分析?自学都可以搞定,学习链接:有参转录组自主分析实操 ;转录组与代谢组结果解读/个性化数据分析 ;
4. 代谢组分析硬件要求不高,个人电脑就可以分析,0基础学习链接:代谢组分析实操
3. 现在才是发表T2T基因组/泛基因组文章的好时机,成本低,好发表,做好高级个性化分析发表高分1区不是梦!T2T基因组/比较基因组/泛基因组分析学习链接:T2T基因组组装与注释分析;动植物泛基因组分析 ;比较基因组分析
4. 群体重测序遗传进化分析+GWAS文章,篇篇10+分,缩短分析周期,提升文章亮点,勇敢冲击NGS顶刊,学习链接:群体遗传进化GWAS分析 ;
5. 传统图位克隆研究方法经久不衰,有遗传群体,有性状,自己就可以做,学习链接:遗传图谱构建与QTL分析
6. 微生物16S/18S/ITS多样性分析和宏基因组分析,学习链接:宏基因组分析 ;微生物16S/18s/ITS多样性分析
7. 细胞器基因组与比较基因组分析是真正的无需实验就可以发表2区期刊的文章思路,成本低,文章好发,性价比极高,学习链接:线粒体基因组分析实操 ;叶绿体基因组分析实操
8. 数据分析硬件利器,个人分析能力的倍增器,好用便宜的云生信服务器,专门为高通量测序数据分析而生,了解链接:云生信服务器
9. 免费好课,个人能力提升,生信入门,不可错过,内容涵盖:生信语言与工具基础、R语言绘图、常见科研绘图、数据处理技能等,免费好课总入口:
10. 更多学习内容:linux、perl、R语言画图,更多免费课程请点击进入组学大讲堂网校学习
- 发表于 2018-04-22 22:49
- 阅读 ( 4511 )
- 分类:转录组
