同源基因概念(直系同源 vs旁系同源 )
分子进化与系统发生
拉马提出进废退理论。他说生物经常使用的器官会逐渐发达,不经常使用的器官会逐渐退化。而且这种后天获得的性状是可以遗传的,因此生物可以把后天锻炼的成果遗传给下一代。达尔文认为,所有的生物物种都存在趋利的适应性变化。并且这些适应性变化会通过一种他称之为“自然选择”的过程遗传下去。而大自然是数百万年间推动演变进化的唯一力量。这个理念让达尔文成为了现代生物学之父。达尔文认为“物竞天择”意味着,存在一种最初的生物,之后通过某种方式得到了改良。
拉马克和达尔文告诉我们,进化很重要!
传统上有两种研究进化的方法。一种是看死的,一种是看活的。看死的,也就是看化石,这是进化最直接,最确凿的证据。第二种研究方法是利用比较形态学、比较解剖学和生理学等手段,确定大致的进化框架。这种方法比第一种方法更容易实现。但是这种方法仅局限于大致的框架,很多细节是存在争议的。这两种传统方法都有局限性,如今研究的是分子进化。也就是利用软件,从分子水平上构建物种的进化树。这里说的分子水平是指 DNA、RNA、以及蛋白质序列。
分子进化理论是 1962 年美国科学家 Linus Pauling提出的。这种理论与传统研究方法的最大区别是,它研究的是 DNA、RNA 以及蛋白质序列这些分子水平上的信息,而不是物种的外在特征。并且基于某一个特定的分子在不同物种中的序列差异来构建系统发生树。此外,分子进化有两个基本的假设条件,只有接受这两个假设,分子进化理论才能得以实施。第一、DNA、RNA 或蛋白质序列包含了物种的所有进化史信息。第二是分子钟理论。这个理论说的是,一个特定基因或蛋白质的进化变异速度在不同物种中是基本恒定的。所谓变异速度是指一定时间内不同碱基或氨基酸突变的个数。这个进化变异速度被认为是恒定的,跟物种没有关系。所以,拿蛋白质来说,两个蛋白质在序列上越相似,他们距离共同祖先就越近。分子钟理论是进化研究领域被普遍认可的理论,但是至今也没有直接的证据证实。
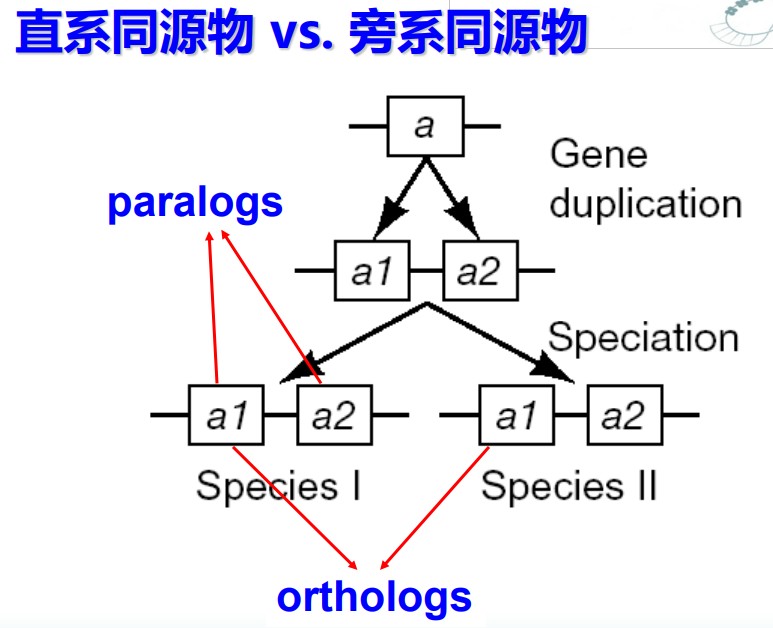
同源(Homologs):来源于共同祖先的相似序列为同源序列。也就是说,相似序列有两种,一种是来源于共同祖先的,那么他们可以叫同源,另一种不是来源于共同祖先的,那么他们尽管相似也不能叫同源。第二种情况出现的概率虽然低,但还是存在的,所以相似序列并不一定是同源序列。同源又分为三种,直系同源,旁系同源和异同源。同源只是对性质的一种判定,只能定性描述,不能定量描述。
直系同源(Orthologs)是指来自于不同物种的由垂直家系,也就是物种形成,进化而来的基因,并且典型的保留与原始基因相同的功能。也就是说,随着进化分支,一个基因进入了不同的物种,并保留了原有功能。这时,不同物种中的这个基因就属于直系同源。
旁系同源(Paralogs)是指在同一物种中的来源于基因复制的基因,可能会进化出新的但与原功能相关的功能来。
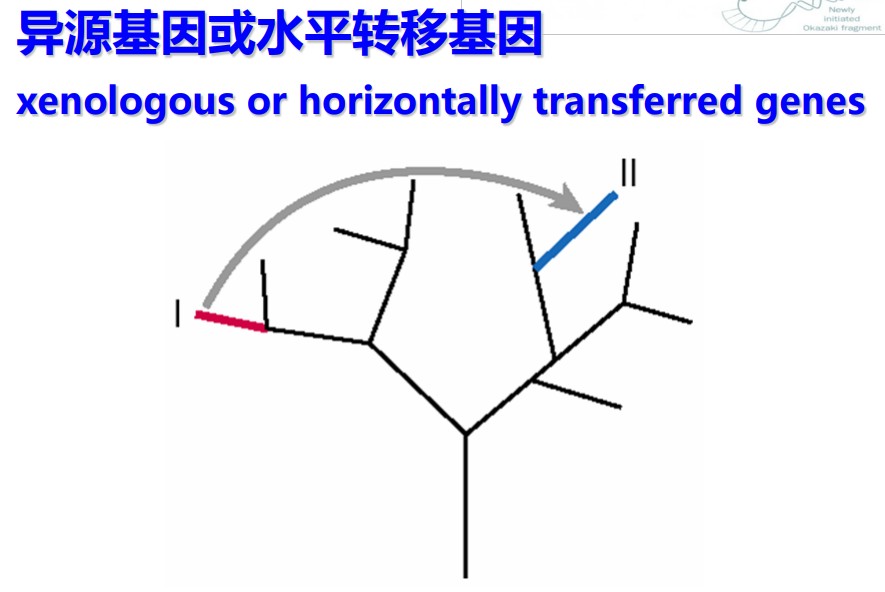
异同源(Xenologs)是指通过水平基因转移,来源于共生或病毒侵染所产生的相似基因。异同源的产生不是垂直进化而来的,也不是平行复制产生的,而是由于原核生物与真核生物的接触,比如病毒感染,在跨度巨大的物种间跳跃转移产生的。
更多生物信息课程:https://study.omicsclass.com/index
- 发表于 2020-09-28 16:37
- 阅读 ( 23910 )
- 分类:遗传进化
