QTL定位方法介绍
QTL(quantitative trait locus),即数量性状位点,是指控制数量性状的基因在基因组中的位置。基于多基因假说的经典数量遗传学通过一些遗传设计和统计模型,把控制某一数量性状的多基因系统作为一个整体来研究,利用一些遗传参数如遗传力和遗传方差来描述数量性状的遗传特征。但是这些参数是所有基因效应的总和,不能有效地分析控制性状表达的基因数目、具体位置、效应大小及其作用方式,因而难以实现数量性状的遗传操纵。遗传标记的发现和使用,促进了数量性状遗传控制研究的发展。人们设想可以把控制数量性状的多个基因分解成单个的QTL,像研究质量性状一样对其分别进行研究。分子标记技术的出现,为深入研究数量性状的遗传基础提供了可能。利用分子标记进行QTL定位原理,实质上就是检测标记与QTL之间的连锁关系,通过计算已知座位的标记与未知座位的QTL之间的交换率来确定QTL的具体位置,同时估计QTL的效应。下面介绍一下QTL定位方法:
1 单标记作图定位法(Single Marker Analysis):
遗传图中的每一个标记按照基因型对表型进行分组看分组之间的均值有没有差异,是否有差异可以用方差分析(检验两个及两个以上样本均数差别的显著性)、t检验、线性回归等方法进行检验,如果有差异说明标记可能与表型连锁,如果没差异表示随机与表型不连锁。如下图所示,每一行是一个样本的基因型与表型数据,每一列为不同的标记,按照A基因型和B基因型进行分组看看Yield的均值在两组之间是否有差异,这就是最简单的单标记关联定位法;
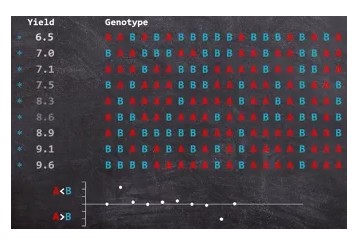
但是单标记作图定位法有缺点,就是无法知道QTL的位置以及计算标记对表型的加显性效应和上位性效应,这里顺便介绍一下这些效应概念:
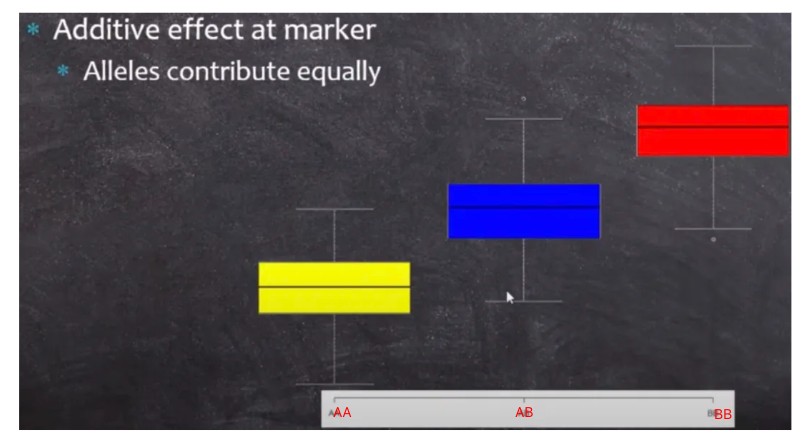
加性效应(additive effect):是指等位基因间和非等位基因间效应的简单相加, 而各个基因对某性状的共同效应就是每个基因对该性状的单独效应的总和。
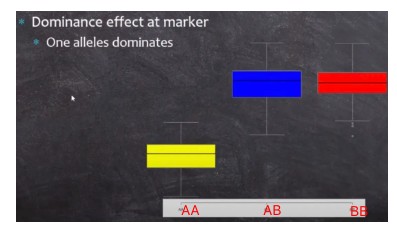
显性效应(Dominance effect):指的是各基因效应值与其加性效应值的离差(deviation)
上位效应(epistasis ):是指不同基因位点的非等位基因之间相互作用所产生的效应。
关于以上效应更多解释说明:https://www.omicsclass.com/article/1616
2 区间作图定位法(interval Mapping):
在线性模型基础上,利用最大似然法对相邻标记构成的区间内任意一点可能存在的QTL进行似然比检验,进而获得其效应的极大似然估计。
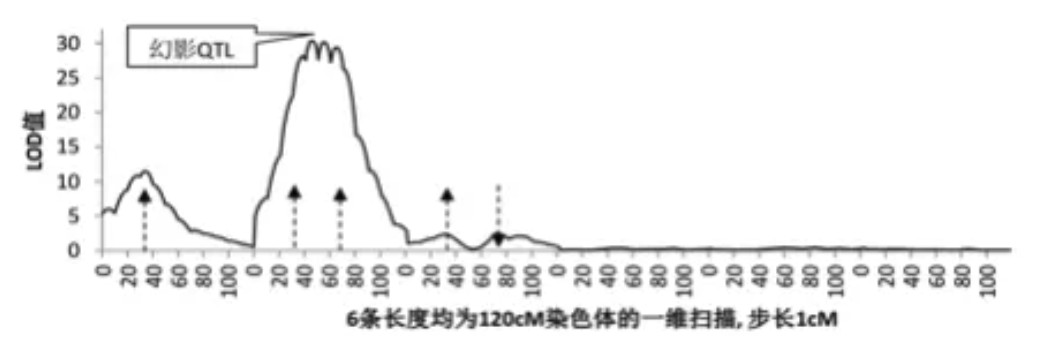
该方法的基本假定是每条染色体上至多包含一个 QTL、QTL的遗传效应满足加—显性模型, 因此当实际情况不符合此假定时, 例如一条染色体上有 2 个 QTL 时, 若两 QTL 的作用方向相反, 往往检测不到; 若作用方向相同, 在两 QTL 间可能会出现一个“幻影”QTL, 且 QTL 位置的置信 区间较大, 一般在 10~30cM 之间, 会造成待估QTL 位置与效应估计值的偏差。
例如下图中:共考虑6条染色体,第一条存在一个QTL,加性效应为1;第二条存在两个QTL,加性效应均为1;第三条存在两个QTL,加性效应为1和-1;其它三条无QTL。图中箭头对应的X轴,为QTL的真实位置,方向表示加性效应的正负。如果存在两个连锁QTL,并且QTL的遗传效应有相 同的方向(又称为相引连锁),区间作图会在两个 QTL的中间出现一个峰,这种现象称为幻影QTL。
3 复合区间作图定位(Composite interval Mapping)
使用逐步回归,将其它与表型相关的QTL作为协变量控制背景遗传效应。通过引入其他标记作为协变量来消除区间以外QTL对作图区间的影响, 从而消除“幻影”QTL 现象, 适用于同一染色体上有多个QTL 的情形。
4 多区间作图法(Multiple Interval Mapping):
多QTL定位的方法主要有极大似 然法和贝叶斯方法两大类, 极大似然法主要包括多 区间作图法和惩罚最大似然法。贝叶斯方法主 要包括可逆跳跃马尔可夫蒙托卡罗(Markov chain Monte Carlo, 简称MCMC)方法[11]和压缩估计方法。多 QTL 同时定位的缺点在于算法太复杂、收敛速度太慢、运算时间太长、而且需要较大的样本量。若标记较多时,难以进行参数估计, 很多贝叶斯模型 无法合理分析一个包含 127 标记、群体大小为145 的大麦 DH 家系,因此, 如何将贝叶斯方法应用于实际作图群体仍有许多问题值得探讨。
5 不同定位法有缺点:

遗传图与QTL定位视频课程推荐:
讲解从高通量测序的SNP数据到遗传图构建以及QTL定位的课程,可点击链接或扫描二维码了解详情:https://bdtcd.xetslk.com/s/4w0i1J

参考资料:
https://iastate.pressbooks.pub/molecularplantbreeding/chapter/cluster-analysis-association-qtl-mapping/
作物学报 ACTA AGRONOMICA SINICA 2009, 35(2): 239−245。DOI: 10.3724/SP.J.1006.2009.00239
- 发表于 2024-06-03 09:44
- 阅读 ( 2725 )
- 分类:遗传图
